 The Architecture of the Connective Tissue in the Musculoskeletal System—An Often Overlooked Functional Parameter as to Proprioception in the Locomotor Apparatus The Architecture of the Connective Tissue in the Musculoskeletal System—An Often Overlooked Functional Parameter as to Proprioception in the Locomotor Apparatus
The Architecture of the Connective Tissue in the Musculoskeletal System—An Often Overlooked Functional Parameter as to Proprioception in the Locomotor Apparatus The Architecture of the Connective Tissue in the Musculoskeletal System—An Often Overlooked Functional Parameter as to Proprioception in the Locomotor ApparatusJaap van der Wal, MD, PhD
University Maastricht, Faculty of Health, Medicine and Life Sciences, Department of Anatomy and Embryology, Maastricht, Netherlands
•••
Editor's Note: This article is based on the doctoral thesis presented by the author at the University Maastricht in 1988: "The Organization of the Substrate of Proprioception in the Elbow Region of the Rat."
•••
The architecture of the connective tissue, including structures such as fasciae, sheaths, and membranes, is more important for understanding functional meaning than is more traditional anatomy, whose anatomical dissection method neglects and denies the continuity of the connective tissue as integrating matrix of the body.
The connective tissue anatomy and architecture exhibits two functional tendencies that are present in all areas of the body in different ways and relationships. In body cavities, the "disconnecting" quality of shaping space enables mobility; between organs and body parts, the "connecting" dimension enables functional mechanical interactions. In the musculoskeletal system, those two features of the connective tissue are also present. They cannot be found by the usual analytic dissection procedures. An architectural description is necessary.
This article uses such a methodologic approach and gives such a description for the lateral elbow region. The result is an alternative architectural view of the anatomic substrate involved in the transmission and conveyance of forces over synovial joints. An architectural description of the muscular and connective tissue organized in series with each other to enable the transmission of forces over these dynamic entities is more appropriate than is the classical concept of "passive" force-guiding structures such as ligaments organized in parallel to actively force-transmitting structures such as muscles with tendons.
The discrimination between so-called joint receptors and muscle receptors is an artificial distinction when function is considered. Mechanoreceptors, also the so-called muscle receptors, are arranged in the context of force circumstances—that is, of the architecture of muscle and connective tissue rather than of the classical anatomic structures such as muscle, capsules, and ligaments. In the lateral cubital region of the rat, a spectrum of mechanosensitive substrate occurs at the transitional areas between regular dense connective tissue layers and the muscle fascicles organized in series with them. This substrate exhibits features of type and location of the mechanosensitive nerve terminals that usually are considered characteristic for "joint receptors" as well as for "muscle receptors."
The receptors for proprioception are concentrated in those areas where tensile stresses are conveyed over the elbow joint. Structures cannot be divided into either joint receptors or muscle receptors when muscular and collagenous connective tissue structures function in series to maintain joint integrity and stability. In vivo, those connective tissue structures are strained during movements of the skeletal parts, those movements in turn being induced and led by tension in muscular tissue. In principle, because of the architecture, receptors can also be stimulated by changes in muscle tension without skeletal movement, or by skeletal movement without change in muscle tension. A mutual relationship exists between structure (and function) of the mechanoreceptors and the architecture of the muscular and regular dense connective tissue. Both are instrumental in the coding of proprioceptive information to the central nervous system.
KEYWORDS: Fascia, dissection, connective tissue, skeletal muscle, proprioception, elbow joint
Some thirty-five years ago, when I received my first training as anatomist, it was not customary to focus one's methodologic attention on the anatomy of connective tissue in general or of fasciae in particular. On the contrary, one was more or less trained to consider fasciae to be connective layers that had to be removed. This approach is related to the fact that the proper method and procedure of anatomy was and still is dissection.
Although dissection is no longer the main approach in visualizing the anatomy and structure of the human body—modern imaging techniques can do so in the living body—"dissectional thinking" still is the main method of analyzing the body in its anatomy. But in the days of my training, one had to separate—to "dissect"— and the revealed structures had to be "cleaned" and "cleared" of connective tissue. Connective tissue was something resembling a covering or sleeve over and between the dissected structures. Therefore it often had to be removed during the dissection procedure.
Most anatomy textbooks today show muscles as discrete anatomic structures with the surrounding and enveloping connective tissue layers removed. When connective tissue was met as a layer, a membrane, a fascia covering a body structure, organ, or region, it was given a name derived from the anatomic substrate that the layer covered. Connective tissue anatomy is often defined as a sub-organization of anatomic structures such as muscles, organs, and so on. Fasciae are thus considered to be "part of" organs and structures.
In leading textbooks, fasciae are therefore defined as "masses of connective tissue large enough to be visible with the unaided eye"(1) (p. 42) and classified as anatomic entities or structures related to organs. But are fasciae, membranes, sheaths in the body in fact distinct and discrete anatomic structures, or are we dealing with continuity? Is the anatomical view missing something when it allocates parts of this fascial continuity to anatomic structures and entities such as body walls or regions (for example, fascia endothoracica or fascia colli media), organs (for example, fascia renalis), or body parts (for example, fascia cruris)? In addition, does a topographic perspective on fascia give any clue about the kind of architectural, functional–mechanical relationship being dealt with?
Schleip mentions the fascia as "the dense irregular connective tissue that surrounds and connects every muscle, even the tiniest myofibril, and every single organ of the body forming continuity throughout the body."(2,3) In this way, fascia is considered an important integrative element in human posture and movement organization (locomotor apparatus) and is often referred to as the "organ of form."(4) Does an analytical and "dissectional" approach to anatomy do justice to this concept?
In removing or dissecting the connective tissue in the form of "layers," every anatomist observes, but often overlooks, various degrees of attachment. Sometimes a layer of fascia is just loosely connected with the underlying or neighboring structure or tissue; sometimes, it is very tight and interwoven with it, and the fascia really has to be cut away, as is the case with the fascia cruris, for example. In both cases, the concept of "dissected means discrete" tends to remain, with fascia viewed as distinct from other tissues, except for those clearly organized in a mechanical in-series relationship with muscular tissue, as in recognized auxiliary structures such as tendons and aponeuroses.
This methodologic mentality has also lead traditional anatomy to dissect the musculoskeletal system into discrete anatomic structures as represented by bones, joints, and muscles. The present article shows that architectural and mechanical spatial relationships between the various tissue components of the musculoskeletal system reveal functional units that go across the traditional anatomic entities of bones, joints, and muscles.
This larger view of functional relationships and coherence is supported by modern neurophysiology. In the central nervous system, the traditional anatomic organization of the musculoskeletal system is only very poorly represented topologically, if at all. The functional and coordinated components of position and motion are not the muscles (and joints), but movements and performed actions. Modern task-dependent models as initiated by Loeb(5,6) indicate that motor units are not necessarily organized in the central nervous system with respect to individual motor nuclei, but according to behavioral tasks. This organization suggests that humans conceptualize a locomotion system in a broader sense, including the coordinating and regulating nervous system (central as well as peripheral), and discriminate that from the locomotion system in the narrower sense (locomotor apparatus), which is represented by the actual musculoskeletal system.
Under the procedural and mental scalpel of the anatomist, the continuity of the connective tissue as central matrix of the body has been lost. The primary connective tissue of the body is the embryonic mesoderm. The mesoderm represents the matrix and environment within which the organs and structures of the body have been differentiated and therefore are embedded. The German embryologist Blechschmidt therefore distinguished the mesoderm as germinal layer: an "inner tissue" in opposition to the ectoderm and endoderm as "limiting tissues." In histology, "limiting tissue" is commonly called epithelium and is constituted almost solely of cells, with relatively little intercellular space. "Inner tissue" could be described as undifferentiated connective tissue, mesenchyme, and is in principle organized in three components: cells, intercellular space (interstitial substances), and fibers.(7,8) Most derivatives of the so-called inner tissue can be identified in histology as connective tissue, including the head-mesenchyme as derivative from neurodermal tissue.
As to the functional development and differentiation of the mesenchyme, there are two patterns of "connection."The first pattern is the development of "intercellular space," which represents a fissure functioning as a sliding and slipping space as is seen in the formation of coelom (body cavities) and of joint "cavities." In this pattern, spatial separation is ensured and therefore motion is enabled. In such cavity formation processes, the primary enlarged intercellular space is lined up and delimited by an epithelium (in body cavities, a so-called mesothelium). Such epithelia more or less depend on the presence of continuous motion for their functional maintenance. Fascial layers such as peritoneum and pleural membrane tend to adhere as soon as the movement of the related structures and organs becomes absent. This phenomenon can also be observed in immobilized joints, showing that, in functional perspective, body cavities and joint spaces have similarities.
The second pattern of development and differentiation of the mesenchyme is the formation of a binding medium, either fibers (as in regular dense connective tissue structures such as membranes and ligaments) or interstitial substrate and matrix (for example, configured in cartilaginous joints). This pattern represents the functional tendency of "connecting" by means of the tissue components of the mesenchyme (one or a combination of cells, intercellular substance, and fibers).
In such a way, a whole spectrum of connectivity could be described in the musculoskeletal system. On the one extreme, connecting structures resemble the desmal sutures in the skull, where dense connective tissue membranes indeed construct a nearly immobile joint connection. The other extreme is represented by the synovial joints (articulations), where the uttermost mobility is exerted. This latter configuration is also shown in the fissures of the body cavities, where organs and body walls and organs themselves are "connected" in a relationship of mobility. The cartilaginous joints (symphyses) more or less represent an intermediate scale of connecting: in humans, nearly all the classical symphyses (such as the ones between the vertebrae or the two pubic bones) tend to the formation of an articulating fissure.
One methodologic restriction has to be made: These concepts are valuable only in a phenomenologic and functional approach. They do not tell anything about the conditions affecting differentiation of these tissues and structures. From the perspective applied here, the primary connective tissue may "connect" ("bind") or it may "dis-connect" ("create room"). Gray's Anatomy states that "joints in principle are connections between bones (arthroses)" but that the "specialized connective tissues of the constituted joints can be either solid or develop a cavity"(1) (p. 103). The synovial joints are called diarthroses. They connect in principle two enchondral bones (with the mandibular and sternoclavicular joints as exceptions). The non-synovial solid joints are called synarthroses. Depending on the properties of the "intervening" connective tissue, the latter are fibrous joints (sutures, gomphoses, and the syndesmoses) or cartilaginous joints (synchondroses). Fibrous joints are usually composed of regular dense connective tissue, sometimes of somewhat more fibroelastic connective tissue.
This view of two types of connectivity is also applicable to the anatomy of fasciae. In general, fasciae in the musculoskeletal system exhibit two different mechanical and functional types:
In osteopathic circles, the continuum and continuity of the "connective tissue apparatus" in the human is emphasized. Such a view is in harmony with the view described here, in particular if the formation of cracks and fissures ("articulating spaces") as a way of "connecting" that enables mobility are considered. The principal function of mesoderm as "inner tissue" is "mediating" in the sense of "connecting" (binding) and "disconnecting" (shaping space). This multiple functionality is reflected in the wavering and divergent classifications that are given to connective tissue in textbooks of anatomy and histology. For example, Gray's Anatomy categorizes connective tissue based on the degree of orientation of the fibrous components: irregular connective tissue (including loose areolar, dense irregular, and adipose tissue) and regular (dense) connective tissue(1) (p. 41). Within the first category, areolar ("loose") connective tissue "holds" organs and epithelia "in place" and has a variety of fibers, including collagen and elastin. Regular dense connective tissue, on the other hand, forms ligaments and tendons. Elsewhere, the book discriminates ordinary ("general") types of connective tissue, special skeletal types (bone and cartilage), and hemolymphoid tissue as a third category(1) (p. 46). The first two in this category are classified as "supportive connective tissue"; bone (osseous tissue) makes up virtually the entire skeleton in adult vertebrates, and in most other vertebrates, cartilage is found primarily in joints, where it provides cushioning.
The usual classifications of connective tissue, including fasciae, not based upon functional criteria are not very consistent in their categories. Gray's Anatomy defines fasciae as "masses of connective tissue large enough to be visible with the unaided eye"(1) (p. 42). As examples of fascia, the sheaths around nerves and vessels are mentioned, as are the fasciae "on the surface" of muscles and organs and between movable muscles, meant as "mechanical isolation." Gray's Anatomy makes special reference to the superficial fascia and to the deep fascia, the latter in particular developed in limbs where it condenses to thicker nonelastic sheaths and cases around the muscles. The discrete anatomic structure (for instance, muscle) is considered as reference, and therefore the fascia is defined as a kind of secondary auxiliary envelope to that (primary) structure. This view of fasciae as a kind of secondary structure actually results from the scalpel of anatomists, who, while cleaning muscles from the fascial layers, have disrupted anatomic continuity where it exists in vivo.
In principle, only two kinds of forces have to be transmitted over synovial joints between the articulating elements in the locomotor apparatus: forces of compression and of tension. Compression forces between the articulating elements are transmitted via the articular surfaces of the adjacent bone elements. The tractive forces and mechanical stresses over the synovial joints are assumed to be transmitted both by passive and by active components in the musculoskeletal system. Regular dense connective tissue structures such as ligaments convey (transmit) those forces "passively."
From here on, the term "connective tissue," if not indicated otherwise, is used in the narrow meaning of "regular dense (collagenous) connective tissue" (RDCT). Such structures can transmit only in a very particular position of the joint—that is, when they are stretched and loaded. That is why this method of force transmission is called "passive." Muscles can transmit force in varying joint positions because they can actively change and adapt their length. Anatomically, the two main components of this system— that is, muscles and ligaments—are generally thought to be organized in parallel. Muscles can control joint stability in the whole range of motion, ligaments only in a particular joint position. This means that the periarticular connective tissue, such as capsules and ligaments, which also play roles in providing mechanoreceptive information to the central nervous system, can be triggered only in a particular joint position—that is, when the relevant connective tissue is stretched or loaded.
Anatomists of the University Maastricht, Netherlands,(9–11) started to study the architecture of RDCT complexes in the musculoskeletal system both globally and specifically in the cubital region. They found that, in the human elbow joint, passive conveyance of tensile stresses does not occur through capsular ligaments alone. In a simulation model of the human elbow joint, the reaction forces resulting from the forces of the biceps and brachioradialis muscle activity and an applied external load showed only a small difference between simulations with an intact capsule (including ligaments) and simulations with the capsule sectioned.(12) These and other findings challenged notions about the mechanical architecture of the periarticular structures in that region and their role in transmitting forces and stresses along the elbow joint. Our team at the University Maastricht therefore developed a new technique of dissection to explore these anatomic relationships.
The central prerequisite in the "alternative" dissection procedure that has been developed was to maintain continuity of the connective tissue by a connective tissue sparing dissection procedure. In the dorsolateral antebrachial and elbow region, the antebrachial fascia (fascia antebrachii) was not removed, but was opened by longitudinal incisions parallel to the long axis of the superficial extensor muscles underneath. The fascia was then released from the muscle fibers of the underlying muscles. In the distal third of the so-called muscle bellies, where the muscle fibers convert to the peripheral tendons of the muscles, this separation from underlying muscle tissue was easily made (Fig. 1). Here, underneath the fascia, a "gliding and sliding" layer of loose areolar tissue was found, similar to tissue in areas of tendinous bursae. Here, the muscles of the dorsal antebrachial region appear as the anatomically separated structures and entities that they are conceptualized to be in anatomy dissections and textbooks. The connective tissue and fascia involved serves as a gliding and sliding—that is, "disconnecting"—medium.
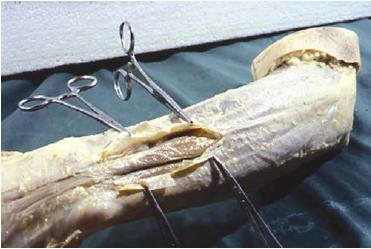
FIG. 1. Opening of the antebrachial fascia in the distal forearm re-gion. Intermuscular loose areolar connective tissue revealed be-tween the discrete muscle bellies and tendons. Left arm, dorsal side, lateral view.
However, in the proximal half of the forearm, the situation is very different (Fig. 2). Here, the muscle fascicles originate from the antebrachial fascia in an oblique or pennate configuration. Only a sharp cutting procedure could "remove" the fascia from the underlying muscle fibers. Those proximal muscle belly fibers were also tightly connected with strong intermuscular connective tissue layers immediately continuous with fascia antebrachii. So, in the proximal lateral cubital, the architecture of the fascial connective tissue is quite different from that in the distal region: a complex apparatus of RDCT layers is situated on top, between, and deep to the muscles. The layers themselves are continuous with each other, forming walls of muscle compartments (cases). The muscle fibers originate in an oblique or pennate configuration from those compartment walls; the walls, in their turn, converge toward the lateral humeral epicondyle. In fact, an epicondylar connective tissue apparatus serves as the insertion area for the neighboring muscle fibers. Tensile forces are therefore transmitted from the muscle fibers to the lateral humeral epicondyle via these converging layers of RDCT. No muscle fiber inserts directly to the epicondyle. Only the most superficial part of the extensor carpi radialis muscle originates from the supracondylar humeral periosteum. In addition, in the proximal lateral cubital region, collagenous fibers do not run from bone to bone as is usually thought. Most of the collagenous fibers in the proximal lateral cubital region appear to be interposed between skeletal tissue and muscle fascicles. Therefore no separate entity such as a collateral radial ligament could be demonstrated.
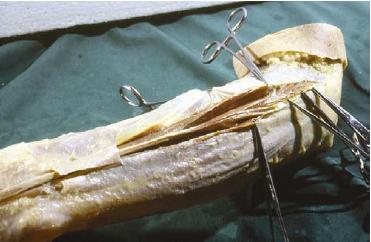
FIG. 2. The compartment walls of the proximal muscle compartment of the third extensor digitorum muscle are opened and separated from the muscle fibers. Left arm, dorsal side, lateral view.
In a "regular" dissection procedure, the next step is that the muscles are dissected and taken out. The scalpel has to cut sharply away the proximal muscle bellies of the extensor muscles, in this way leaving in situ strong bands of collagenous connective tissue that could be identified as collateral radial ligament. In the connective-tissue sparing dissection, the muscle fibers are removed and the already-mentioned epicondylar connective tissue apparatus is revealed. The RDCT strands that are usually identified as collateral ligament are indeed an integral part of the epicondylar connective tissue apparatus, meaning that, in regular dissection, the collateral radial ligament is dissected out as an artifact! This is demonstrated in Figs. 3 and 4.
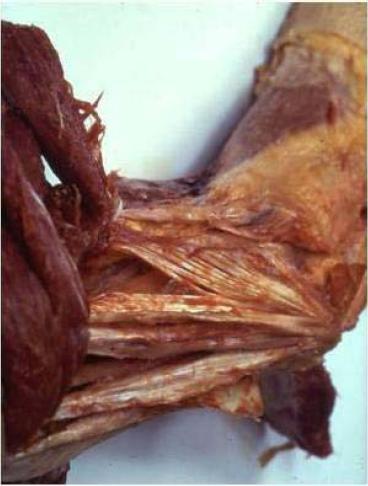
FIG. 3. Proximal lateral elbow region. Muscles are dissected away from the epicondylar connective tissue apparatus and reflected (to the left). The convergence of the remaining connective tissue muscle compartment walls toward the lateral humeral epicondyle is clearly demonstrated. Left elbow, lateral view.
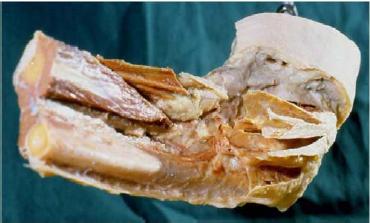
FIG. 4. Proximal lateral forearm region. Muscles and muscular tis-sue have been removed. The most proximal extensions of the muscle compartment walls (the epicondylar connective tissue apparatus) are left in situ, demonstrating the muscle compartments converging to the lateral epicondyle. Left elbow, lateral view.
The same situation appears to be true for the annular radial ligament. The proximal portion of the supinator muscle appears as a broad and long aponeurotic structure. This aponeurosis merges with the other layers and is an integral part of the epicondylar connective tissue apparatus converging to the lateral humeral epicondyle. Not any muscle fiber of the supinator muscle has a bony insertion on the humeral epicondyle itself.
Again, when the supinator muscle is dissected as an "entity," a strand of collagenous connective tissue remains that might be identified as annular ligament. However, the collagenous fibers of this band run in proximodistal direction and not in a circumradial direction as is usually represented in anatomy books. If an annular radial ligament is dissected, it will exhibit cut edges, indicating that, again, a mechanical continuity has been distorted in the effort to dissect ligaments and muscles as parallel structures. In fact the so-called annular ligament does not exist; it is an integrated part of the aponeurotic layer of connective tissue via which supinator muscle fibers insert to the lateral epicondyle. This layer in turn is an integrated part of an epicondylar connective tissue apparatus.
The proximal lateral cubital region holds a complex apparatus of RDCT layers that mainly consist of muscle compartment walls that converge toward the lateral humeral epicondyle. Only a single muscle, the anconeus, inserts directly into the humeral periosteum as seen earlier with the extensor carpi radialis. However, most collagenous fibers in the proximal lateral cubital region are interposed between skeletal tissue and muscle fascicles. Only a very small portion of the fibers run from bone to bone and may therefore be classified as ligamentous fibers. Indeed no separate entities such as collateral or annular ligaments can be described. This means that most muscle fibers in the proximal lateral elbow region are organized in series and not in parallel with the connective tissue of this apparatus. The muscle/connective tissue units form the functional units that transmit tensile stresses over the elbow joint, with muscular and collagenous connective tissue organized in series. These units do not coincide with the usual anatomic classification into muscles and ligaments. Although such functional units do indeed coincide with muscles and their distal tendons as both functional and morphologic entities in the distal extent of the forearm, the functional organization is seen to be transmuscular (or "non-muscular") in the proximal forearm region.
This architecture has consequences for conveying tensile forces and stresses over a synovial joint. Usually it is assumed that two components in the musculoskeletal system convey tensile mechanical stresses over synovial joints: CT structures such as ligaments convey such forces passively, and muscles serve as the "active" components, the latter structures organized in parallel to the former ones. Ligaments can perform their force-conveying function only in a very particular position of the articulating bones—that is, they must be stretched and loaded. On the other hand, muscles are capable of this function in varying positions of the joint, because they are able to continuously adapt in length. Here, this is called the in-parallel view, and it is demonstrated in Fig. 5(a,b).
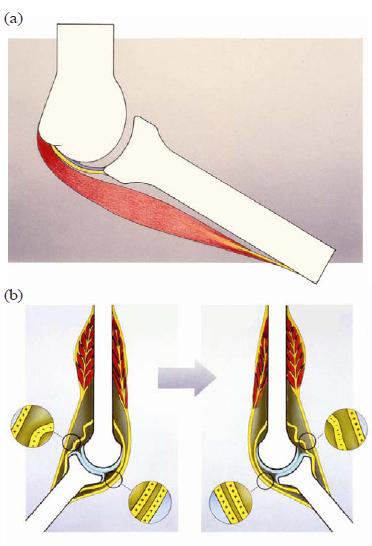
FIG. 5. (a) The "classical" in-parallel organization of the iuxtaarticular tissue. From inside to outside: articular capsule (blue); reinforcing iuxta-articular regular dense connective tissue structures (ligaments) (yellow); and on the outer side, periarticular muscle (red). (b) The "classical" organization principle of iuxta-articular connective tissue running from bone to bone, organized in parallel to the muscular component (tendons). Only in a particular joint position can the connective tissue transmit forces or signal in the sense of mechanoreceptor triggering (++++ versus ––––).
In an in-series configuration as alternatively described here, the conveying of tensile stresses by the collagenous fibers also depends on the muscle fascicles that are active. In vivo displacement of bones and muscular activity influence the state of stress and tension of connective tissue elements. In this model, passive and active joint-stabilizing structures organized in parallel—that is, muscles and ligaments with mutually independent functions—cannot be distinguished. The joint capsule and its reinforcements no longer have an exclusive role in the passive conveying of tensile stresses. The functional units involved in the transmission of forces do not consist of topographically defined and separate entities of either muscular or ligamentous tissue.
For instance, a structure such as the supinator aponeurosis may be classified as epimysial fascia, but also as an aponeurosis or even as a "ligament" with adjustable length and tension: a "dynament" (discussed in more detail shortly). The traditional topographic approach to the locomotor apparatus also assumes that the passive components (ligaments) are deep to the superficial components (muscles) that are actively involved in the maintenance of joint stability and integrity [Fig. 5(a,b)]. This concept is challenged by the in-series architecture described here. In the lateral cubital region of humans and rats, no ligaments can be distinguished as separate entities. There is one joint stability system, in which muscular tissue and RDCT interweave and function mainly in an in series situation as shown in Fig. 6(a,b). Thus, in vivo, the periarticular connective tissue is loaded and stretched both by the movement of related skeletal parts and by the tension of the muscle tissue inserting to this connective tissue.
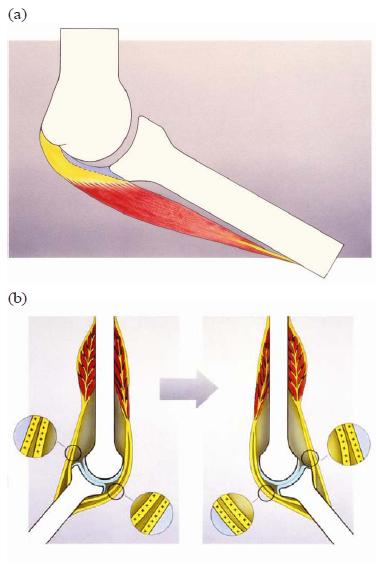
FIG. 6. (a) The alternative in-series organization of the iuxta-articular tissue. From inside to outside: articular capsule (blue); periarticular regular dense connective tissue (yellow) in series with periarticular muscle (red). (b) The alternative organization of iuxtaarticular connective tissue organized in series to the muscular component. In all joint positions the connective tissue of the joint is brought to tension and is capable of transmitting forces and signaling in the sense of mechanoreceptor triggering (++++ and ++++).
This connective tissue architecture can be better appreciated if, rather than talking in terms of collateral ligaments, a "lateral cubital force transmission system" (LCFTS) is defined(9) that can be made visible in a magnetic resonance imaging section of the region. This approach reveals a principle that can be recognized in many other areas and regions of the body. For example, similar connective tissue architecture has been described for the opposite region of the elbow: a "medial cubital force transmission system." The in-series continuity of the patellar retinacula (including parts of the so-called collateral ligaments of the knee joint) with the vastus medialis and vastus lateralis muscles demonstrates the same principle. Similar architectural relationships are seen for the fascia cruris and the ventral extensor muscles of the foreleg, or for the erector spinae muscle and the thoracolumbar fascia.
Detailed studies of the lateral cubital region of the rat (discussed later in this article) showed this architecture.(10) Most deep and superficial RDCT layers (as muscle compartment walls) are organized in series with muscle fascicles. Collagenous fibers running from bone to bone—thought to be stressed passively by displacement of the articulating bones—hardly occur. Instead, there occur broad aponeurotic layers of RDCT to which relatively short muscle fascicles insert, which, on the opposite side, are directly attached to skeletal elements. Such configurations of muscle fascicles attached to the periosteum of one articulating bone and via a layer of RDCT indirectly attached to another articulating bone, could be considered "dynamic ligaments." Such "dynaments" are not necessarily situated directly beside the joint cavity or in the deep part of the joint region.
By describing the dynament as an architectural unit of the musculoskeletal system, we mean a unit of RDCT connected to the periosteum of a skeletal element with muscle fascicles in series attached to it. In Fig. 7(a) a dynament is represented in its most basic appearance: a unipennate muscle between two skeletal elements. A typical unipennate forearm muscle as represented in Fig. 7(b) shows the common appearance of the dynament. In this situation, the distal RDCT element is represented by the tendon and is situated intramuscularly. The proximal RDCT element is represented by one of the muscle compartment walls and is situated extramuscularly.
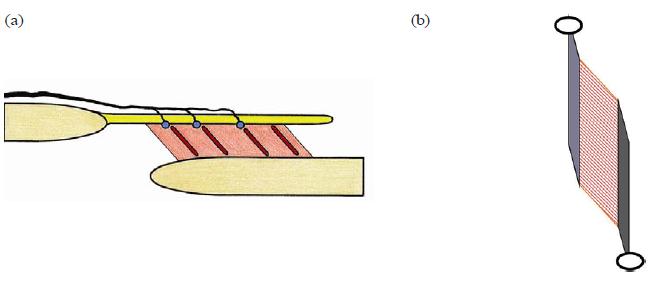
FIG. 7. (a) Schematic diagram of the "dynament" as architectural unit. A regular dense (collagenous) connective tissue (RDCT) layer (top, yellow) with inserted muscle portion (middle, red). Morphologic substrate of proprioception indicated with blue dots (LC, RC–GTO, see text) and red stripes (muscle spindles). Afferent nerve indicated (on top, black). Note that the innervation pattern of the muscle-related mechanoreceptors resembles the innervation pattern of a joint capsule (from outside to inside). (b) An unipennate forearm muscle as typical "dynament." Proximal (top left, light grey), an RDCT layer (membrane, aponeurosis, septum, etc.) with muscle fascicles attached to it, mostly extramuscular (middle, striated red). Distal (bottom right, dark grey), an RDCT layer (tendon, aponeurosis, etc.) with muscle fascicles (middle, striated) attached to it, mostly intramuscular. In this basic situation, the muscle component is organized as intermediate between two RCDT structures.
Distal forearm extensor carpi radialis brevis muscles and extensor digitorum muscles clearly show this type of functional unit [Fig. 8(c)]. The supinator muscle shows a variant configuration, with distal connective tissue layers that are well developed, but without an extensive connective tissue apparatus intermediating at the insertion [Fig. 8(b)]. The long head of the triceps shows a similar orientation but in the "opposite direction" [Fig. 8(d)], with developed connective tissue layers proximally rather than distally. If neither a "proximal" nor a "distal" connective tissue apparatus has been developed, the result is a muscle "without aponeurosis or tendon"—for example, the deltoideus muscle [Fig. 8(a)]. If the muscular connecting and intermediating tissue has completely "disappeared," a ligament is the consequence [Fig. 8(e)].
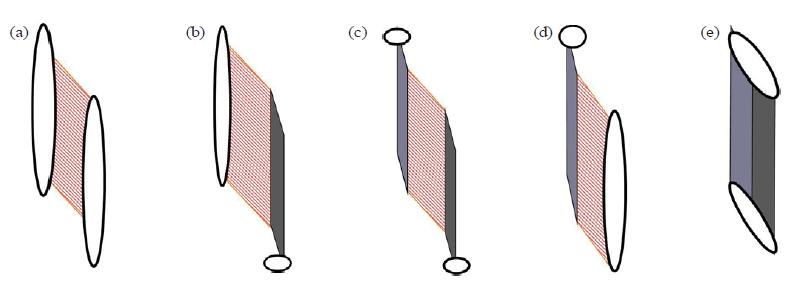
FIG. 8. (a,b,c,d,e) Possible appearances of the "dynament" as architectural unit. In the middle (c), the basic situation [see Fig. 7(b)]. On the left (b), muscle tissue proximally inserting directly to the skeletal element (periosteum) and distally via tendons. On the right (d), muscle tissue distally inserting directly to the skeletal element (periosteum), proximally via septa and aponeuroses. On the extreme left (a), only muscular fascicles, no intermediating regular dense connective tissue (RDCT) structure—a "typical muscle." On the extreme right (e), no muscle tissue intermediating, only RDCT—a "typical ligament."
In fact, ligaments, defined as strands of RDCT running from the one skeletal element to the next, are an exception. They have to be an exception. The RDCT has tissue properties of high resistance to loading, a high degree of hysteresis, and little elasticity; a ligament as interposed structure between two movable bones can therefore be constructed only if the distance between the opposite points of ligamentous insertion on the bones changes very little during the range of motion of the joint. This requires specialized design in the configuration of the bones and the joint (surfaces), and there are only a few examples of such "true" ligaments at joints in the body: cruciate ligaments of the knee joint or ligamentum apicis dentis in the atlantooccipital joint.
Of course in the "classical" fibrous joints, the ligamentous organization principle is clearly present, but in such cases, this organization is consistent with the way such joints are functionally loaded. Such configurations could therefore be considered extremes to the general rule. It bears repeating that these phenomenologic and functional considerations do not tell anything about the conditions in and by which those tissues and structures differentiate.
An architectural approach to the anatomy of the locomotion system as practiced here shows that fasciae exhibit a variety of mechanical relationships with neighboring tissue and therefore may play quite different functional roles. Sometimes they act as aponeuroses, sometimes they are gliding envelopes building joint-like gliding spaces. The nomenclature "fascia" should therefore be considered and reevaluated critically in every region. The "classical" fasciae of the organs and of muscles usually represent the "gliding fasciae" type (again, we consider the coelom as a kind of "joint space"). Many epimysial muscle fasciae function in a similar way. However, a fascia such as the fascia cruris or the retinaculum patellae functions as an epimuscular aponeurosis.
To understand the mechanical and functional circumstances for the fascial role in connecting and conveying stresses, it is more important to know the architecture of the connective and muscle tissue than the regular anatomical order or topography. In principle, this approach applies to every fascial layer in the human body. One must know both where they are situated ("anatomy") and how they are connecting and connected ("architecture").
The architectural view developed here has implications for the understanding and interpretation of the spatial organization of the nervous afferents, with related receptors that form the substrate for proprioception. What kind of role does the architecture in the musculoskeletal system play in the quality of centripetal information from the various components and tissues of the system?
The usual distinction between muscle afferents and articular afferents is implicitly based on the anatomical concept that (peri)articular RDCT structures and muscular tissue structures are organized in parallel to each other along the joint. This latter concept is the one that was challenged in the previous part of this essay.
Connective tissue and fasciae are richly innervated.(3,2) Fascial layers may thus play an important role in proprioception and nociception. Considerations such as "architecture versus anatomy (topography)," mutatis mutandis may also apply for the spatial organization of mechanoreceptors, the morphologic substrate for proprioception. To study the role and function of mechanoreceptors in the process of proprioception, it may be important to know where they actually are located in such regions and how they are or are not connected with the relating tissue elements. The actual spatial organization of such receptors can be better interpreted functionally when it is known how their topography is related to the architecture of the connective and muscular tissue.
Proprioception is the process of conscious and subconscious sensoring of joint position or motion. Encapsulated or unencapsulated mechanosensitive sensory nerve endings (mechanoreceptors) and related afferent neurons provide the centripetal information needed for the control of locomotion or for the maintenance of posture. In general, such mechanoreceptors are reported to occur either as muscle receptors or as joint receptors. Muscle receptors are mechanoreceptors present in the muscles, including their auxiliary structures such as tendons, aponeuroses, and fasciae. Muscle spindles and Golgi tendon organs (GTOs) are the best-known types of such receptors.(13) Joint receptors are considered to be situated in joint capsules and related structures, including reinforcing ligaments. These receptor types are usually ordered according to the (ultra)structure of the receptor itself, physiologic features, type of afferent nerve fiber, and other parameters.(14–17,a)
In the traditional view, joint receptors play the leading role in monitoring joint position or movement for statesthesis and kinesthesis; muscle receptors are relegated to motor functions that operate at a subconscious or reflex level (reviewed by McCloskey(18) and Matthews(19)). However, this concept has been challenged by physiology investigations suggesting that muscle spindle afferents can also contribute to human kinesthesis. Clinical observations in patients who retained their kinesthesis after complete surgical removal or interruption of joint capsules (endoprostheses) and experiments that tested position-sensing abilities following selective anesthesia of joint capsules in vivo provided further evidence that muscle spindles (and mechanoreceptors of the skin) also contribute to kinesthesis.(20–23) It is often stated that joint receptors react only at the extremes of joint position, acting as "limit detectors".(24) Indeed, it has been found that the discharge of articular receptors is limited to positions that significantly stress the joint capsule—positions in which the capsule and related structures are supposed to be strained passively.(18,25–28) Other studies suggest that muscle afferents provide the substrate of mid-or full-range receptor activity (or both) present in recordings from articular nerves.(18,26–31) In their well-known studies, Abrahams, Richmond, and Bakker describe in the neck region of the cat a variety of sensory endings in so-called non-articular connective tissue. In neck proprioception, they suggest attributing a more important role of mechanoreception to substrate that is not situated directly within or near joint capsules.(32–34)
Mechanoreceptors are in fact free nerve endings (FNEs), whether or not equipped with specialized end organs. The main stimulus for such receptors is deformation. Variation exists as to the microarchitecture of the ending. On the one hand, there exists the principle of lamellae around a relatively simple nerve ending. This represents the principle of the ball- or bean-shaped Vater Paccini or paciniform corpuscles, often called lamellated corpuscles (LC). On the other hand, there is the more spray-like organization of the nerve ending wrapping around and between the deformable substrate-like connective tissue fibers. Those are the spindle-shaped Ruffini corpuscles (RC) or GTOs. These two types of microarchitecture roughly relate to the type of mechanical deformation that is at stake—that is, compression for the lamellated bodies and traction and torsion for the spray-like type. Other varying parameters are threshold, adaptivity, and adjustability. In this general classification, the muscle spindle is a spindle-shaped spray-like ending organized around specialized muscle fibers equipped with the extra possibility of adjustable length.
Mechanoreceptors associated with muscles, including the muscle auxiliary structures such as tendons, are usually classified(14–17) as follows:
The mechanoreceptors typically associated with joints are these:
The assumption of an in-series organization of muscular and collagenous connective tissue instead of an in-parallel organization strongly influences the view of the areas that may be considered to be most "strategic" for mechanoreception based on deformation or other mechanical stimuli. Deformation is expected to be highest in the transition between tissues of the musculoskeletal system with different consistencies and mechanical qualities. Similar considerations hold for the transitional areas of muscular and collagenous connective tissue with skeletal tissue.
Twenty-five years ago, our research group at Maastricht University proposed this question: Is there a correlation between the architecture of the muscular and connective tissue and the spatial distribution ("architecture") of the morphologic substrate of proprioception?
In the rat, innervation of the RDCT apparatus in the proximal lateral elbow region was studied using various techniques and reconstructed three-dimensionally in relation to the architecture of muscle and connective tissue.(10,11) The findings from those studies showed that no mechanosensitive nerve terminals are present within the stress-conveying components of the epicondylar connective tissue apparatus. However, FNEs are found in the transitional areas of the connective tissue apparatus towards the periosteum. Particularly in the outer (epimysial) and in some intermuscular components of the apparatus, plexiform-arranged nerve fascicles can be demonstrated. Besides autonomic nerve fibers, they contain group III (A-delta) and group IV (C–) nerve fibers. Those nerve fibers are involved in the afferent pathway of proprioceptive information from the transitional areas between the connective tissue layers and the muscle fascicles organized in series with them [shown schematically in Fig. 7(a)]. This also seems to represent a more ligamentous or articular "pattern of innervation" compared with the related nerve fascicles running on the "outside" of the innervated structure. This is actually a typical capsular or articular pattern [see Fig. 7(a)]. Such plexuses are also very dominantly present on the so-called ante-brachial fascia. The nerve fascicles of such substrate terminate in LCs, RCs, GTOs, FNEs, and (sometimes) muscle spindles.
A spectrum of mechanosensitive substrate occurs at the transitional areas between the RDCT layers and the muscle fascicles organized in series with them. This substrate exhibits features of the mechanosensitive nerve terminals that usually are considered to be characteristic for "joint receptors" and for "muscle receptors." For example, the olecranal retinaculum and the supinator aponeurosis (and its proximal continuation with the epicondylar connective tissue apparatus) with the muscle fascicles in series to those layers are well-equipped with such morphologic substrate. The human equivalents of these structures are, respectively, the aponeurosis of the supinator muscle and the tendon of the triceps muscle.
Based on architecture and spatial distribution of the substrate of mechanoreception, it may be assumed that the "joint receptors" here are also influenced by the activity of the muscle organized in series with the collagenous connective tissue near those receptors. The proximal extensions of the muscle compartment walls are well-equipped with mechanosensitive substrate. This supports the idea that the stresses during joint positioning are conveyed mainly via those collagenous layers and also are involved in triggering the related mechanoreceptors. In the studied region, there exists no basis in morphology for so-called joint receptors that are deformed exclusively by passive strain in collagenous connective tissue structures induced by displacement of the articulating bones.
The demonstrated substrate of proprioception also caused trouble as to morphologic classification. The typology and spectrum of nerve fascicles and nerve terminals found here exhibit features usually associated with tendons as well as with joint capsules and ligaments.
Summarizing, then, we have encountered
The substrate of proprioception that we found in and near the RDCT apparatus has features of mechanoreceptors that usually are linked with "joint receptor" substrate and with the mechanoreceptors usually present in muscles and related tendons.
The findings regarding spatial distribution and organization of the mechanoreceptive substrate that is usually accepted as muscle receptors—that is, the muscle spindles and GTOs—were even more suitable with the concept that is brought forward here. The latter receptors are not organized according to principles of anatomy and topography, but according to the functional architecture of RDCT layers in relation to muscular architecture. In Fig. 9(a,b), it is clearly shown that, in all the studied antebrachial extensor muscles, the distribution of muscle spindles per muscle area is uneven. If the spatial distribution of muscle spindles is considered per muscle, it is difficult to detect a common distribution pattern in all muscles. But considering muscle spindle and GTO distribution "transmuscularly" reveals other functional aspects of the distribution pattern. The spatial distribution and orientation of the spindles (including the distribution of GTOs) is understandable only from the regional functional architecture of the RDCT structures. The muscular zones that are dense in muscle spindles and GTOs are the stress- and force-conveying zones of the muscle that are in series with the LCFTS distally and in series with the peripheral tendons proximally. This arrangement provides a common principle that may explain many kinds of distribution patterns. And, of course, sometimes architectural units coincide with specific topographic entities.
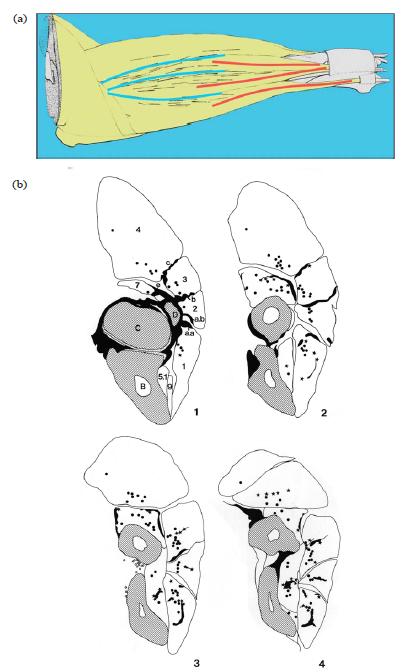
FIG. 9 (a) The spatial distribution of muscle spindles in the superficial lateral forearm muscle in the rat. The distribution is clearly more related
to the architecture of the proximal epicondylar connective tissue apparatus than to the topography of the muscles. The spindles are presented as thin black lines. The thicker lines in the diagram represent the intermuscular septa that are part of the proximal regular dense connective tissue (RDCT) apparatus (blue, on the left) and the distal tendons of the superficial extensor muscles (red, on the right). (b) Typical cross-sections of a rat forearm (four proximal sections from a total of six forearm sections). The muscle spindles and Golgi tendon organs (GTOs) in a given segment are projected onto each section in a summative projection. Dots are muscle spindles, stars are GTOs. Note that GTOs are not only present near or at distal tendons, the proximal intermuscular septa and fasciae also have GTOs arranged to it.
We have demonstrated that muscle spindles and GTOs occur in muscular tissue areas in series with RDCT structures. Such an area does not often coincide with a morphologic muscle entity. The architectural relation between the RDCT structures seems to define the spatial distribution of spindles and GTOs in the region as a whole. We identified (sub)units of muscular tissue in series with RDCT, so-called muscle spindle/GTO zones, with a relatively high density of mechanoreceptors. The patterns of distribution and orientation in such zones are understandable from the perspective of the architecture of the muscular tissue related to the connective tissue architecture.
Muscle spindles and GTOs are concentrated mostly in areas of muscular tissue directly intermediating distal and proximal RDCT structures—that is, tendons distally, compartment walls proximally. If distal and proximal RDCT structures are situated at a relatively large distance from each other, the receptors accordingly form small elongated zones segregated from surrounding extrafusal fibers (see Fig. 10, patterns 1 and 2). If the distance between distal and proximal RDCT structures is relatively short, both structures are situated more in parallel with each other: the spindles run obliquely in parallel to each other in a relatively broad muscular area bridging a short distance. Individual GTOs occur in a direct one-to-one relation to individual spindles in a relatively broad area of muscle/connective tissue transition. The extrafusal fibers are more or less equally distributed between them (see Fig. 10, patterns 3 and 4). Muscle portions that do not link two RDCT structures, but that attach directly to periosteal tissue, tend to be devoid of muscle spindles.
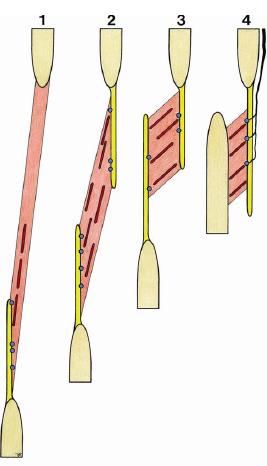
FIG. 10. Typical patterns for muscle spindle/Golgi tendon organ moni-tor zones (see text). The configuration as shown at 4 (right) repre-sents the typical pattern of a “dynament.” [See also Fig. 7(a)].
An in series unit of muscular tissue/RDCT layer/ skeletal element equipped with mechanosensitive substrate at the transitional areas between the various tissue components constitutes the basic unit of the spatial organization of the substrate of proprioception. Such a unit may occur as a muscle fraction in series with a muscle compartment wall that is shared with the muscular tissue of an adjacent muscle. It may also appear as a muscle compartment wall with muscle fascicles inserting unilaterally and with afferent nerve fibers reaching the related mechanoreceptors from the outer side. This was introduced earlier as the typical "dynamic ligament" (dynament—see Fig. 10, pattern 4).
If the number of mechanoreceptors is calculated (per weight or per volume), the outcome depends not only on the number of receptors, but also on the extension, volume, or magnitude of the unit that is taken into account. This finding is again determined by how the entities in the locomotor apparatus are conceived of. In the model proposed here, neither individual muscles nor ligaments are the functional entities to which receptor distribution in the relevant joint region should be related.
For example, muscles with similar quantitative densities appeared to exhibit completely different distribution patterns, and those with similar distribution patterns showed different densities. Similar considerations are valid both for absolute and for relative receptor volumes, because quantifying receptors per muscle or per connective tissue structure ignores the functional architectural continuity in the various tissue elements of the locomotor apparatus that maintains joint integrity. Quantitative parameters of mechanoreceptors fail to express the functionality of their spatial distribution related to the architecture of muscle and connective tissue. For example, muscle spindle density may make more sense reported by a (joint) region or a (peri)articular area than by muscle.
Another direct consequence of the concept proposed here is that the organization of the morphologic substrate of proprioception should be regarded in terms of fractions of muscular tissue rather than in terms of muscles. Recent research suggests that, also on the level of spinal sensorimotor control, muscles should no longer be considered the functional entity in the locomotor system.(35–38) In addition, the organization principle of neuromuscular muscle compartments projecting in a topographical organization to the corresponding motor nucleus is thought to allow the organism to differentiate muscle activity into activity of muscle parts. This concept again matches well the task-dependent model of Loeb: that is, that motor units are not necessarily organized with respect to individual motor nuclei, but according to behavioral tasks. The concept of the locomotor apparatus being built up by architectural units of muscular tissue in series with collagenous connective tissue is more consistent with such trans- or supramuscular models than is the concept in which muscles function as the entities that maintain joint integrity.
The muscle spindles and GTOs in the lateral cubital region of the rat are concentrated in those areas where (in view of the description of the architecture of the muscle and tissue) the conveying of tensile stresses over the elbow joint is expected to take place. So the spatial organization of muscle spindles and GTOs in the studied region is such that it enables monitoring of the stresses conveyed over the elbow joint and of the movements of the articulating bones. This organization allows those receptors to be classified in this situation also as "joint receptors." The spatial organization shows the principle that mechanoreceptors are arranged in tissue environments that "offer" them deformation. In these environments, the spatial architecture of the connective tissue apparatus is a predominant factor.
In consequence of the identification of an in-series organization of muscular tissue and RDCT (mainly tendons distally and compartment walls proximally) attached to skeletal parts (periosteal attachment), three configuration types of mechanoreceptors can be identified:
In the above-mentioned configurations, RCs are not indicated as a separate category. The GTOs and RCs are considered to be the same receptor type, presenting gradual differences depending on the texture of the surrounding tissue.(45,47,48) It may therefore be stated that the quartet MS–GTO/RC–LC–FNE represents the complete spectrum of mechanoreceptors in a joint region. In this way, the three main types of so-called muscle receptors—MS, GTO, and LC(13)—are combined with the three types of so-called capsular (or joint) receptors—RC, LC, and FNE.(43)
The conclusion is that, in vivo, the activity of a mechanoreceptor is defined not only by its functional properties, but also by its architectural environment. If Abrahams, Richmond, and Bakker(34) state that the topography of mechanoreceptors provides a "subtle comparative function in the process of sensory coding of muscle events," they raise the important issue of the spatial distribution of receptors in the process of proprioception. To this should be added the notion that the architecture of the muscular and connective tissue and consequent receptor distribution plays a significant role in the coding of the proprioceptive information that is provided.
I wish to thank Professor Thomas W. Findley, MD, PhD, of the Center for Healthcare Knowledge, VA Medical Center, East Orange and Newark NJ, USA, for his extensive effort to read, comment, and edit this article thoroughly and precisely. A part of the intelligibility of this article is a result of his tremendous effort to convert my Dutch English to good scientific language. This expression of gratitude also regards his assistant, Mr. Thomas Brown. Figures 1 through 6 are used courtesy of the Department of Anatomy and Embryology, University of Maastricht, The Netherlands.
The author declares that there are no conflicts of interest.
Published under the CreativeCommons Attribution NonCommercial-NoDerivs 3.0 License .
Corresponding author: Jaap van der Wal, University Maastricht, Faculty of Health, Medicine and Life Sciences, Department of Anatomy and Embryology, PO Box 616, Maastricht 6200 MD Netherlands.
E-mail: walembryo@home.uni-one.nl
Footnotes:
a O'Connor and Gonzales being based on the work of Freeman and Wyke.
(Return to Text)
INTERNATIONAL JOURNAL OF THERAPEUTIC MASSAGE AND BODYWORK—VOLUME 2, NUMBER 4, DECEMBER 2009